Mistrz Programowania
Omówienie zadania Dąb Bajtek
Zanim zacznę omawiać zadanie, przypomnę definicję ocalonego wierzchołka, ponieważ będziemy z niej często korzystać. Na końcu dnia wierchołki połączone z jednynką, nie odciętymi krawędziami są ocalone. W grupie tych wierzchołków nie może znaleźć się żaden zainfekowany.
Ponieważ zadanie jest całkiem skomplikowane, zanim przejdziemy do tego w jaki sposób poruszać się po drzewie (czyli docelowego rozwiązania), postaramy się uprościć treść jak tylko możliwe.
Obserwacja 1
Nigdy nie ocalimy wierzchołka $v$, który należy do poddrzewa pewnego zainfekowanego wierzchołka $u$.
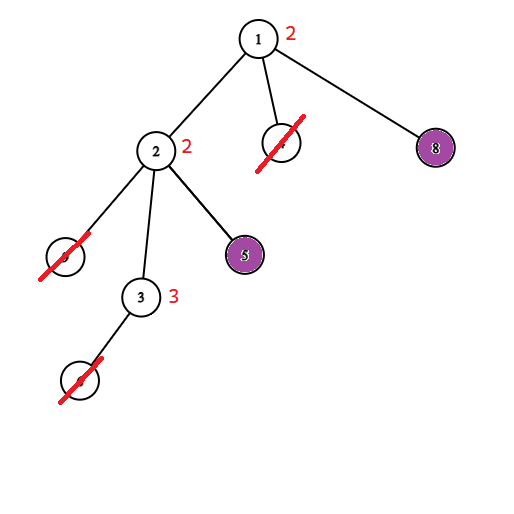
Dowód: Jeżeli $v$ byłby ocalony, to byłby połączony z jedynką nie odciętymi krawędziami, a ponieważ ten graf jest drzewem, to takie połączenie jest dokładnie jedno i przechodzi przez $u$. Zatem $u$ też byłby połączony z jedynką, co jest niedopuszczalene, bo $u$ jest zainfekowany. Możemy więc pozbyć się wszystkich poddrzew zainfekowanych wierzchołków, i zostaniemy z drzewem, w którym każdy zainfekowany wierzchołek jest liściem (jak na rysunkach poniżej).
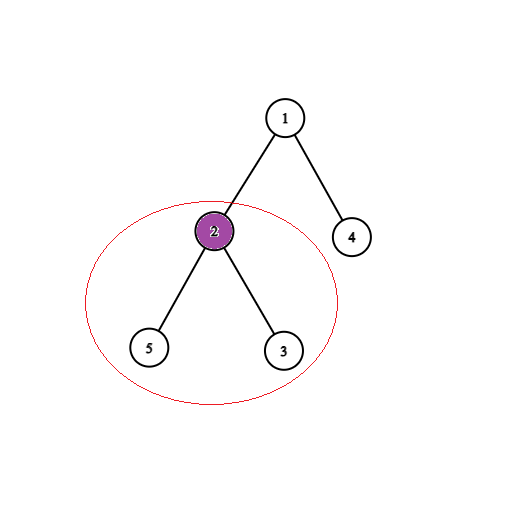
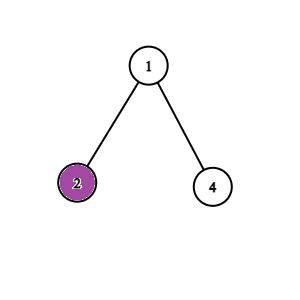
Obserwacja 2
Przyjrzyjmy się teraz liściom, które nie są zainfekowane. Niech $v$ będzie zdrowym liściem, a $u$ jego ojcem. Zauważmy, że jeżeli ocalimy $u$, to bez żadnej straty możemy też ocalić $v$.
Dowód: Wystarczy nie ucinać krawędzi $v \leftrightarrow u$, skoro $u$ był ocalony, to $v$ wtedy też będzie oaclony i nie połączymy w ten sposób żadnego zainfekowanego wierzchołka z jedynką, bo jedyny wierzchołek jaki dodamy to $v$.
Możemy więc znowu skompresować nasze drzewo. Możemy usunąć wierzchołek $v$ całkowicie i zapisać w wierzchołku $u$ informację (wagę), że jeżeli go ocalimy, to dodamy do wyniku o jedgen wierzchołek ($v$) więcej. Potem możemy powtarzać ten zabieg dopóki wszystkie liście nie będą zainfekowane. Poniżej przykład takiego procesu.
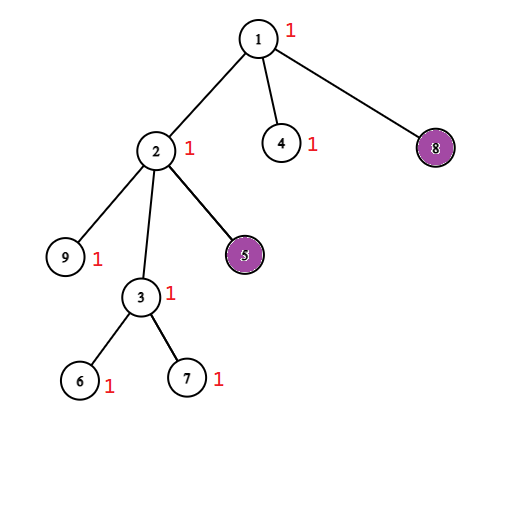
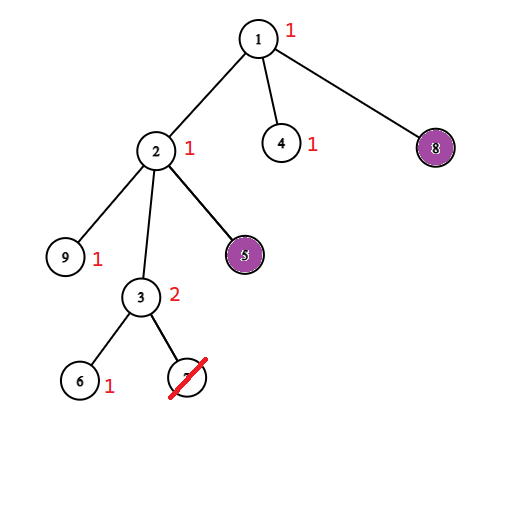
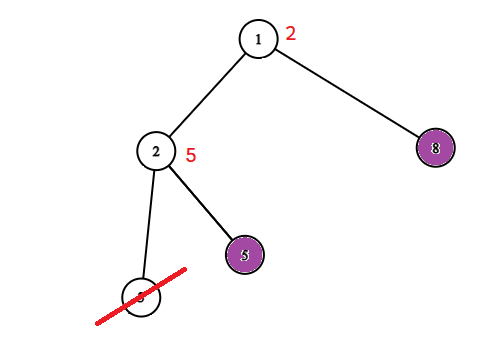
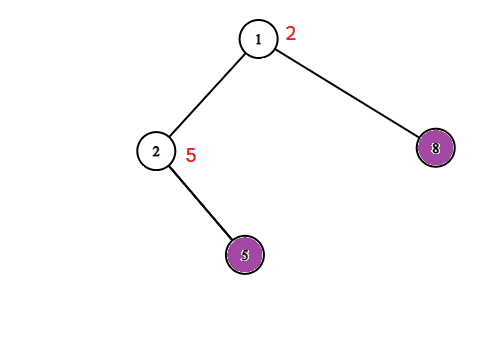
Po zastosowaniu tych dwóch kompresji zostaniemy z drzewem o ważonych wierzchołkach, w którym wszytkie liście są zainfekowane oraz wszystkie pozostałe wierzchołki są zdrowe. Waga wierzchołka w tym drzewie mówi nam ile powinnniśmy dodać do wyniku ocalając dany wierzchołek. 1
Od teraz mówiąc “wierzchołek” będę miał na myśli niezainfekowany wierzchołek w tym skompresowanym, ważonym drzewie.
Obserwacja 3
Możemy ocalić wszystkie wierzchołki które odwiedzimy.
Dowód: Zaczynamy w jedynce, więc jedynkę zawsze odwiedzimy. Możemy się przemieszczać na 2 sposoby, albo przejście wzdłuż krawędzi, albo powrót do jedynki (która jest odwiedzona). Zatem wszystkie odwiedzone wierzchołki będą połączone ze sobą, a więc również z jedynką. Nie będzie wśród nich żadnych zainfekowanych, a skoro każdy z nich jest w pewnym momencie odwiedzony, to możemy odciąć wszystkie krawędzie łączące tą grupę z nie odwiedzonymi wierzchołkami.
Obserwacja 4
Nie możemy ocalić żadnego wierzchołka którego nie odwiedzimy.
Dowód: Załóżmy że chcemy ocalić nie odwiedzony wierzchołek $v$ ($v \neq 1$, bo $1$ jest zawsze odwiedzony). Ponieważ nigdy go nie odwiedzimy, to w szczególności nigdy nie będziemy też w jego poddrzewie, zatem nigdy nie utniemy tam żadnych krawędzi. Musielibyśmy więc ocalić całe to poddrzewo. W każdym poddrzewie musi być przynajmniej jeden liść, a ponieważ każdy liść jest zainfekowany, to musielibyśmy ocalić zainfekowany wierzchołek, sprzeczność.
Czyli sprowadziliśmy tą treść do takiej postaci: “Dane jest drzewo ukorzenione w 1, ważone na wierzchołkach. Dozwolone są 2 operacje, przejście wzdłuż krawędzi (o koszcie 1) oraz powrót do korzenia (o koszcie 0). Chcesz odwiedzić wierzchołki o możliwie największej sumie wag, zaczynająć w 1 i wydając co najwyżej $t$ jednostek czasu.” 2
że ocalimy dokładnie te wierzchołki które odwiedzimy.
Rozwiążemy ten problem programowaniem dynamicznym.
Policzymy tablicę $DP[v][t][stay]$ która będzie nam mówiła jaką maksymalną sumę możemy uzyskać z poddrzewa wierzchołka $v$, zakładając że zaczynamy w wierzchołku $v$ i mamy do dyspozycji $t$ czasu. $stay$ jest boolem, jeżeli ma wartość $true$, to możemy “zostać” w tym poddrzewie, czyli nie interesuje nas gdzie się znajdujemy po upływie czasu $t$. W przeciwnym wypadku nie możemy “zostać” w tym poddrzewie i chcemy po upływie czasu $t$ znaleźć się spowrotem w wierzchołku $v$. Poniżej przykład.
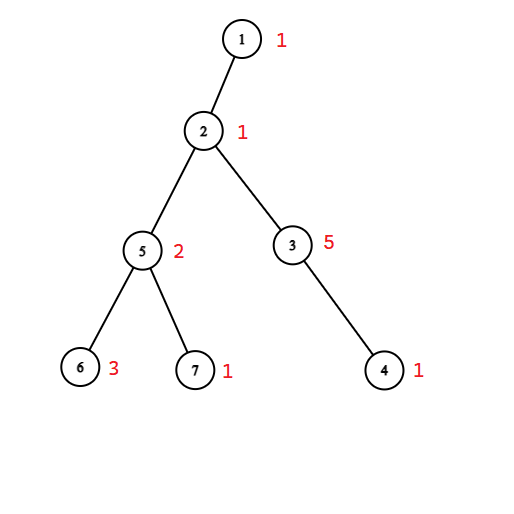
W powyższym grafie $DP[2][7][true] = 13$, ponieważ w 7 jednostek czasu możemy odwiedzić całe poddrzewo 2 w taki sposób: $2 \rightarrow 5 \rightarrow 6 \rightarrow 5 \rightarrow 7 \Rightarrow 1 \rightarrow 2 \rightarrow 3 \rightarrow 4$, gdzie pojedyncze strzałki oznaczają przejście krawędzią (koszt 1), a podwójne skok do 1 (koszt 0). Zauważmy, że w tym rozwiązaniu odwiedziliśmy też jedynkę, ale nie dodajemy jej do wyniku, bo nie należy do poddrzewa 2. Jeżeli natomiast zmienimy wartość $stay$ na $false$, to $DP[2][7][false] = 12$ i możemy ten wynik osiągnąć w taki sposób: $2 \rightarrow 5 \rightarrow 6 \rightarrow 5 \rightarrow 7 \Rightarrow 1 \rightarrow 2 \rightarrow 3 \rightarrow 2$.
Jeżeli udałoby nam się policzyć taką tablicę w sensownym czasie, to wynikiem będzie $DP[1][t][true] = DP[1][t][false]$.
Rozważmy najpierw liście. Niech $w[v]$ będzie wagą wierzchołka $v$, a $l$ liściem, wtedy: $DP[l][t][stay] = w[l]$ niezależnie od $t$ oraz $stay$, ponieważ $DP$ zakłada że zaczynamy w $l$, więc $l$ będzie zawsze odwiedzony.
Rozważmy treaz wierzchołek $v$ nie będący liściem. Wzór na $DP[v][t][stay]$ nie jest tak prosty do zapisania, więc tylko wspomnę jak go policzyć. Dla każdego syna $s$ wierzchołka $v$ rozważymy jedną z 3 opcji.
- Nie wchodzimy do $s$.
- Wchodzimy do $s$ (kosztem 1), spędzamy $t’$ czasu w $s$ i wracamy spowrotem do $s$ (dodajemy do wyniku $DP[s][t’][false]$) i wracamy spowrotem do $v$ (kosztem 1). W sumie kosztuje nas to $t’ + 2$ czasu, i dodamy do wyniku $DP[s][t’][false]$.
- Wchodzimy do $s$ (kosztem 1), spędzamy $t’$ czasu w $s$ (dodajemy do wyniku $DP[s][t’][true]$) i zostajemy w tym poddrzewie. W sumie kosztuje nas to $t’ + 1$ czasu, i dodamy do wyniku $DP[s][t’][true]$, ale na końcu zostaniemy w poddrzewie $s$.
Sprawdzając wszystkie możliwości podzielenia $t$ na synów $v$, pamiętając o tym że z opcji 3 może skorzystać tylko ostatni syn, jeżeli liczymy $DP[v][t][true]$, lub żaden jeżeli liczymy $DP[v][t][false]$, otrzymamy poprawne, ale wolne rozwiązanie. Można je jednak znacznie przyspieszyć stosując 2 optymalizacje.
Optymalizacja 1
Zamiast rozważać wszystkie możliwe podziały $t$ na synów $v$, będziemy rozpatrywać tych synów po kolei. Niech $DP_i[v][t][stay]$ będzie wartością $DP[v][t][stay]$, zakładając możemy wchodzić tylko do $i$ pierwszych synów $v$ (niech $s_i$ będzie $i-tym$ synem $v$). 3
Dla $i = 0$ wygląda to identycznie jak dla liścia, natomiast dla pozostałych $i$, aby policzyć $DP_i[v][t][stay]$ przeiterujemy się po $t’$, spróbujemy spędzić $t’$ czasu w pierwszych $i-1$ synach $v$ (co odczytamy za pomocą policzonego wcześniej $DP_{i-1}[v][t][stay]$), a pozostałe $t - t’$ spędzimy w $i-tym$ synie (co odczytami z policzonego wcześniej $DP[s_i][t][stay]$). 4
Optymalizacja 1 daje nam już złożoność $O(n\cdot t^2)$, ponieważ dla każdego wierzchołka $v$ który nie jest 1 ani pierwszym synem innego wierzchołka i każdego $t$ przeiterujemy się po wszystkich $t’$ aby policzyć $DP$ dla ojca $v$.
Optymalizacja 2
Nie musimy dla każdego wierzchołka sprawdzać wszystkich możliwych $t$. W szczególności niezależnie od struktury drzewa, dla $t$ rzędu dwukrotności rozmiaru poddrzewa da się odwiedzić całe drzewo i wrócić spowrotem do wierzchołka startowego.
Przy poprawnej implementacji Optymalizacji 2 złożoność amortyzuje się do $O(n \cdot min(n, t))$.
Dowód: https://ceoi2017.acm.si/files/ceoi2017-solutions-practice.pdf – omówienie zadania Muzeum, na samym dole po “Further optimization”.
Uwagi
- Warto wspomnieć, że przy kompresji drzewa nigdy nie usuwamy wierzchołków które nie są liściami, więc odległości pomiędzy wierzchołkami pozostają równe 1.
-
Przypadek szczególny: Nie ma żadnych zainfekowanych wierzchołków i całe drzewo zostanie skompresowane do jednego wierzchołka, wtedy technicznie jest to nie zainfekowany liść, ale nie będziemy rozważać tej możliwości dalej w rozwiązaniu, bo jest prosta do “wyifowania” ↩
-
Pozbyliśmy się zainfekowanych liści z drzewa, ponieważ nie są potrzebne do rozwiązania, wiemy ↩
-
Celowo nie podałem $i$ jako kolejnego wymiaru $DP$, ponieważ nie chcemy tego fizycznie pamiętać w kodzie ze względu na pamięć. Wartości będziemy trzymać bezpośrednio w $DP[v][t][stay]$, będą się one poprostu zmieniać wraz ze wzrostem $i$. ↩
-
Uwaga: niekoniecznie w tej kolejności. Możemy najpierw pójść do $s_i$, a potem wrócić do poprzednich $i-1$ synów, żeby na końcu zostać w jednym z nich. ↩